Đại Cương Về Kháng Thể
 |
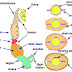
| Cấu trúc điển hình của một kháng thể. |
Các kháng thể này sẽ lưu hành trong máu và bạch huyết, chúng hoạt động như những thành phần thực hiện của đáp ứng miễn dịch dịch thể bằng cách phát hiện và trung hoà hoặc loại bỏ các kháng nguyên. Các kháng thể thực hiện hai chức năng chính: chúng gắn một cách đặc hiệu vào một kháng nguyên và chúng tham gia trong một số chức năng sinh học khác. Chương này chúng ta sẽ tập trung vào vấn đề cấu trúc của các phân tử kháng thể. Đặc điểm cấu trúc nào đã tạo ra tính đặc hiệu và chức năng của những phân tử này.
1. Cấu trúc cơ bản của kháng thể
Từ đầu thế kỷ XX người ta đã biết rằng các kháng thể là các phân tử thực hiện của đáp ứng miễn dịch dịch thể xuất hiện trong huyết thanh. Năm 1939 Tiselius .A và Kabat .E .A đã tiến hành phát hiện phần protein huyết thanh chứa các kháng thể. Các tác giả đã gây miễn dịch cho thỏ bằng ovalbumin sau đó chia huyết thanh của thỏ đã được gây miễn dịch thành hai phần bằng nhau. Phần thứ nhất được diện di để phân tách thành bốn thành phần: albumin và các a-, b- và g- globulin; Phần thứ hai được cho phản ứng với kháng nguyên để hình thành chất kết tủa, sau đó loại bỏ chất tủa này rồi sử dụng phần huyết thanh còn lại để điện di. So sánh hình ảnh điện di của hai mẫu huyết thanh này, các tác giả phát hiện thấy trong mẫu điện di thứ hai các thành phần trong phần g-globulin giảm đi đáng kể. Như vậy phần g-globulin có chứa các kháng thể của huyết thanh và vì vậy chúng được đặt tên là globulin miễn dịch để phân biệt với các protein khác còn lại trong phần g-globulin.
Vào những năm 1950 - 1960 Porter .R và Edelman .G đã chứng minh cấu trúc cơ bản của globulin miễn dịch. Công trình này có một ý nghĩa đáng kể và hai nhà khoa học đã được trao giải thưởng Nobel về y học năm 1972. Các cách tiếp cận thực nghiệm của Edelman và của Porter rất khác nhau. Porter phân cắt các phân tử globulin miễn dịch bằng các enzyme để thu được các mảnh peptide, trong khi đó Edelman đã tách phân tử globulin miễn dịch bằng cách khử các cầu disulfide liên chuỗi. Kết quả của mỗi phương pháp đã bổ sung cho nhau và cho phép chúng ta hiểu được cấu trúc cơ bản của phân tử globulin miễn dịch. Cả Porter và Edelman lần đầu tiên đã phân tách phần g-globulin của huyết thanh bằng siêu ly tâm để thu được hai phần nhỏ: phần thứ nhất có trọng lượng phân tử cao có hằng số lắng là 19S; phần thứ hai có trọng lượng phân tử thấp và hằng số lắng là 7S. Các tác giả đã tách riêng phần 7S xác định được trọng lượng phân tử là 150.000 Da và ký hiệu là globulin miễn dịch G (IgG). Porter đã phân cắt IgG bằng enzyme papain thành các mảnh khác nhau. Mặc dù papain là một enzyme hoạt động thuỷ phân protein không đặc hiệu và phân cắt toàn bộ phân tử protein, nhưng nếu thời gian tác dụng ngắn thì enzyme này chỉ phân cắt các cầu disulfide nhậy cảm nhất. Trong điều kiện như vậy papain đã phân cắt phân tử IgG thành hai mảnh giống nhau (mỗi mảnh có trọng lượng phân tử là 45.000) được gọi là các mảnh Fab (antigen binding fragment), bởi vì mảnh Fab vẫn giữ nguyên khả năng gắn với kháng nguyên của phân tử kháng thể nguyên vẹn. Ngoài hai mảnh Fab tác giả còn thu được một mảnh nữa có trọng lượng phân tử là 50.000 được gọi là mảnh Fc vì khi bảo quản trong lạnh chúng bị kết tinh hoá (chữ c bắt nguồn từ chữ cristalliable nghĩa là có khả năng kết tinh). Nisonoff .A cũng tiếp cận nghiên cứu bằng cách tương tự nhưng dùng enzyme pepsin thay cho enzyme papain. Khi cho pepsin tác dụng ngắn với phân tử IgG thì thu được một mảnh có trọng lượng phân tử 100.000 gồm 2 mảnh Fab gộp lại và ký hiệu là F(ab')2. Mảnh F(ab')2 cũng gây kết tủa kháng nguyên. Tuy nhiên khi xử lý bằng pepsin không thu được mảnh Fc mà thay vào đó là một số mảnh peptide nhỏ.
Cấu trúc chuỗi của IgG lần đầu tiên được nghiên cứu bởi Edelman và sau này được khẳng định bởi Porter. Edelman đã khử các cầu disulfide của phân tử IgG bằng mercaptoethanol rồi tiến hành điện di trên gel tinh bột trong môi trường urê 8M để khử các cầu disulfide trong chuỗi và liên chuỗi làm cho phân tử được trải ra mà không gấp. Tác giả thu được 2 vệt điện di và điều này chứng tỏ rằng phân tử IgG có nhiều hơn một chuỗi protein. Porter cũng phát triển thí nghiệm này bằng cách tạo ra phản ứng khử nhẹ hơn sao cho chỉ cắt các cầu disulfide liên chuỗi mà thôi, sau đó tiến hành alkyl hoá các nhóm sulfhydryl (SH) lộ ra bên ngoài bằng iodoacetamide để ngăn cản sự tái tạo ngẫu nhiên của các cầu disulfide. Ngoài ra tác giả còn cho thêm acid propionic hữu cơ để ngăn cản sự ngưng tập. Sau đó tiến hành sắc ký trên cột để để phân tách các phân tử dựa trên kích thước của chúng (hình 5-2b). Thí nghiệm này đã cho thấy phân tử IgG có trọng lượng phân tử 150.000 Da được hợp thành bởi hai chuỗi peptide, mỗi chuỗi có trọng lượng phân tử 50.000 được ký hiệu là các chuỗi nặng (chuỗi H - heavy chain) và hai chuỗi mỗi chuỗi có trọng lượng phân tử là 25.000 được ký hiệu là các chuỗi nhẹ (chuỗi L - light chain).
Vấn đề còn lại là đi xác định xem các sản phẩm phân cắt bởi enzyme, Fab, F(ab’)2 và Fc có liên quan như thế nào với các sản phẩm phân rã của chuỗi nặng và chuỗi nhẹ? Để trả lời câu hỏi này Porter đã sử dụng kháng huyết thanh dê được gây miễn dịch bởi các mảnh Fab và Fc của phân tử IgG của thỏ. Ông đã phát hiện thấy rằng kháng thể kháng Fab có thể tương tác với cả chuỗi nặng và chuỗi nhẹ, trong khi đó kháng thể kháng Fc thì chỉ phản ứng với chuỗi nặng mà thôi. Điều này chứng tỏ Fab có chứa các thành phần của cả chuỗi nặng và chuỗi nhẹ còn Fc thì chỉ chứa các thành phần của chuỗi nặng. Các kết quả này đã khẳng định mô hình đầu tiên về cấu trúc phân tử IgG mà Porter đã đưa ra. Theo mô hình này thì phân tử IgG bao gồm hai chuỗi nặng giống hệt nhau và hai chuỗi nhẹ giống hệt nhau được liên kết với nhau bằng các cầu disulfua. Enzyme papain phân cắt ngay phía trên cầu disulfua liên chuỗi nối giữa hai chuỗi nặng còn enzyme pepsin thì phân cắt ngay phía dưới cầu disulfua này và vì thế hai enzyme thuỷ phân protein tạo ra các sản phẩn phân rã khác nhau. Khi sử dụng mercaptoethanol để khử và akyl hoá sẽ cho phép ta tách riêng được các chuỗi nặng và chuỗi nhẹ.
2. Các quyết định kháng nguyên trên phân tử globulin miễn dịch
Do các kháng thể là các phân tử glycoprotein nên bản thân chúng cũng có thể hoạt động như một chất sinh miễn dịch để kích thích sinh ra một đáp ứng tạo kháng thể. Người ta chia các quyết định kháng nguyên (hoặc các epitop) trên phân tử globulin miễn dịch thành ba loại chính: các quyết định isotyp, các quyết định allotype và các quyết định idiotyp. Các quyết định này được định vị trên các vị trí đặc trưng của phân tử kháng thể.
2.1. Các quyết định isotype
Các quyết định isotype nằm trên vùng hằng định cho phép phân biệt các chuỗi nặng của các lớp và các lớp nhỏ kháng thể khác nhau, và phân biệt các týp và các týp nhỏ của các chuỗi nhẹ trong một loài nhất định. Mỗi một quyết định isotype được mã hoá bởi một gene của vùng hằng định riêng biệt và tất cả các cá thể trong cùng một loài đều có cùng các gene vùng hằng định mã hoá các quyết định này. Trong cùng một loài, từng cá thể sẽ biểu hiện tất cả các quyết định isotype trong huyết thanh của chúng. Các loài khác nhau được di truyền các gene của vùng hằng định khác nhau và bởi vậy chúng có các isotype khác nhau. Do đó khi lấy globulin miễn dịch của loài này tiêm vào loài khác thì các quyết định isotype sẽ được nhận diện như là lạ và kích thích sinh ra đáp ứng tạo kháng thể chống lại các quyết định isotype của phân tử globulin miễn dịch lạ. Các kháng thể kháng isotype thường được sử dụng để xác định các lớp hoặc các lớp nhỏ của globulin miễn dịch trong huyết thanh hoặc xác định lớp kháng thể gắn trên bề mặt tế bào B (SIg).
2.2. Các quyết định allotyp
Mặc dù tất cả các cá thể trong cùng một loài thừa hưởng cùng một loại bộ gene isotype nhưng lại có một số gene allele khác nhau giữa các cá thể. Những gene allele này mã hoá một số đoạn peptide có trình tự các acid amine khác nhau, người ta gọi đó là các quyết định allotype. Các quyết định này chỉ xuất hiện ở một số chứ không phải là tất cả các thành viên của một loài. ở người đã xác định được allotype của 4 lớp nhỏ IgG, 1 lớp nhỏ IgA và chuỗi nhẹ. Các quyết định allotype của chuỗi (được ký hiệu là Gm. Cho đến nay đã nhận dạng được 25 loại quyết định allotype Gm, chúng được ký hiệu bằng chữ G (viết tắt của chữ gamma) sau đó là số thứ tự lớp nhỏ rồi đến số thứ tự allotype [ví dụ G1m(1), G2m(23), G3m(11), G4m(4a)]. Người ta chỉ mới xác định được allotype của lớp nhỏ IgA2 được ký hiệu là A2m(1) và A2m(2). Chuỗi nhẹ (có 3 quyết định allotype được ký hiệu là (m(1), (m(2), và (m(3). Mỗi allotype này có sự khác nhau từ 1 đến 4 acid amine được mã hoá bởi các gene allele khác nhau.
Các kháng thể chống các quyết định allotype có thể thu được bằng cách tiêm các globulin miễn dịch của một cá thể này cho một cá thể khác trong cùng loài không có các quyết định allotype tương ứng. Kháng thể kháng allotype cũng có thể xuất hiện ở người phụ nữ trong quá trình mang thai khi đáp ứng với quyết định allotype của người chồng có trên phân tử globulin miễn dịch của người con. Kháng thể kháng allotype cũng có thể xuất hiện sau khi truyền máu.
2.3. Các quyết định idiotype
Trình tự các acid amine có duy nhất ở vùng VH và VL của một phân tử kháng thể biết trước có thể hoạt động không chỉ như một vị trí kết hợp kháng nguyên mà còn hoạt động như một quyết định kháng nguyên. Các quyết định idiotype được sinh ra do các vùng thay đổi của chuỗi nặng và chuỗi nhẹ hình thành một cấu trúc đặc biệt về không gian. Mỗi một kháng nguyên riêng lẻ của vùng thay đổi được xem như là một quyết định idiotype hay còn gọi là idiotope.Trong một số trường hợp idiotope có thể là một vị trí kết hợp kháng nguyên thực sự, nhưng trong một số trường hợp một idiotope cũng có thể là một đoạn của vùng thay đổi nằm ngoài vị trí kết hợp kháng nguyên. Mỗi phân tử kháng thể sẽ có nhiều idiotope. Tất cả các idiotop của một phân tử kháng thể sẽ tạo ra hình ảnh idiotype của kháng thể đó.
Bởi vì các kháng thể do các tế bào B nằm trong cùng một clone sản xuất ra có vùng thay đổi giống hệt nhau nên chúng có cùng một hình ảnh idiotype. Các kháng thể không cùng một clone tế bào B sản xuất ra cũng có thể có quyết định idiotype giống nhau. Quyết định idiotype này được gọi là quyết định idiotype chung giống nhau, chúng phản ảnh sự huy động phổ biến cùng một gene vùng thay đổi của tế bào mầm bởi các tế bào B khác nhau của cùng một dòng thuần chủng. Các kháng thể kháng idiotype được tạo ra bằng cách thu hẹp đến mức tối thiểu sự khác biệt isotype hoặc allotype để đến mức có thể nhận dạng được sự khác biệt idiotype. Thông thường có thể sử dụng một kháng thể thuần khiết như protein u tuỷ hoặc một kháng thể đơn clone. Tiêm kháng thể này cho một cơ thể nhận đồng gene thì sẽ hình thành được kháng thể kháng idiotype.
Một trong những thí nghiệm sớm nhất để chứng minh sự có mặt của các quyết định idiotype là thí nghiệm của Oudin .J và Michel .M tại viện Pasteur Paris năm 1963. Các tác giả này đã tiêm kháng thể của thỏ kháng Samonella vào một con thỏ khác cùng nhóm allotype. Thỏ được tiêm đã sản sinh ra kháng thể có khả năng kết hợp được với kháng thể đã dùng để gây miễn dịch mà không kết hợp với những kháng thể của chính con thỏ đó kháng lại các kháng nguyên khác và cũng không kết hợp với kháng thể của những con thỏ khác kháng lại Samonella. Kháng thể kháng idiotype còn được sinh ra bởi động vật trong quá trình đáp ứng miễn dịch và đóng vai trò quan trọng trong việc điều hoà đáp ứng miễn dịch (xem những phần sau).
3. Các lớp kháng thể
Mỗi lớp kháng thể được phân biệt bởi sự khác nhau về trình tự các acid amine trong vùng hằng định chuỗi nặng.
3.1. Kháng thể IgG
IgG có nồng độ cao nhất trong huyết thanh, chiếm khoảng 80% tổng lượng globulin miễn dịch trong huyết thanh. Phân tử IgG là một monomer gồm có hai chuỗi nặng ( và hai chuỗi nhẹ hoặc ( hoặc (. Trọng lượng phân tử của IgG khoảng 150.000 Da, hằng số lắng 7S, vì vậy IgG có thể thấy cả ở trong lòng mạch và ở ngoài lòng mạch. Có 4 lớp nhỏ IgG ở người được đánh dấu theo nồng độ giảm dần của chúng trong huyết thanh: IgG1 (9 mg/ml), IgG2 (3 mg/ml) IgG3 (1 mg/ml), và IgG4 (0,5 mg/ml). Bốn lớp nhỏ này được mã hoá bởi 4 gene vùng hằng định chuỗi nặng (gene CH) khác nhau mà có 90 - 95% trình tự ADN giống nhau. Đặc trưng cấu trúc để phân biệt lớp nhỏ này với lớp nhỏ khác là kích thước của vùng bản lề, số lượng cũng như vị trí của các cầu disulfide liên chuỗi nối các chuỗi nặng (hình 5.12). Sự khác biệt về acid amine giữa các lớp nhỏ của IgG đã làm cho hoạt tính sinh học của phân tử có những khác nhau. IgG1, IgG3 và IgG4 có thể chuyển vận dễ dàng qua nhau thai và đóng vai trò trong việc bảo vệ thai phát triển. Một số lớp nhỏ IgG có thể hoạt hoá bổ thể mặc dù hiệu quả của chúng khác nhau. Lớp nhỏ IgG3 hoạt hoá bổ thể hiệu quả nhất, tiếp theo là IgG1 rồi đến IgG2 còn IgG4 thì không có khả năng hoạt hoá bổ thể. IgG cũng hoạt động như một kháng thể opsonin do chúng có thể gắn vào thụ thể dành cho Fc có trên bề mặt đại thực bào, nhưng chức năng này cũng thay đổi tuỳ theo lớp nhỏ: IgG1 và IgG3 có ái lực cao với thụ thể dành cho Fc, trong khi IgG4 có ái lực yếu hơn và IgG2 có ái lực rất yếu.
3.2. Kháng thể IgM
IgM chiếm 5 - 10% tổng lượng globulin miễn dịch huyết thanh, có nồng độ khoảng 1 mg/ml. IgM monomer xuất hiện ở trên bề mặt tế bào B (SIgM). Loại IgM này được phát hiện ở trên bề mặt của 90% số tế bào B trong máu ngoại vi và có vai trò sinh học như một thụ thể dành cho kháng nguyên. IgM do tế bào plasma tiết ra có cấu tạo pentamer do 5 đơn vị monomer nối với nhau bởi các cầu disulfide giữa các lãnh vực của đầu C tận cùng chuỗi nặng . 5 đơn vị monomer này bố trí sao cho phần Fc quay về phía trung tâm của pentamer và 10 vị trí kết hợp kháng nguyên quay ra phía ngoại vi của pentamer. Mỗi một pentamer có thêm một chuỗi polypeptide được gọi là chuỗi J. Chuỗi J có vai trò trong quá trình polymer hoá các monomer để hình thành pentamer. Chuỗi J được gắn với các gốc cystein ở đầu C tận cùng của 2 trong số 10 chuỗi nặng bằng cầu disulfide.
IgM là lớp globulin miễn dịch đầu tiên xuất hiện trong đáp ứng lần đầu với một kháng nguyên và cũng là lớp globulin miễn dịch đầu tiên được tổng hợp ở trẻ sơ sinh. Cấu trúc pentamer của IgM đã làm cho lớp kháng thể này có một số tính chất riêng biệt. Hoá trị của phân tử tăng lên vì chúng có 10 vị trí kết hợp kháng nguyên. Một phân tử IgM có thể gắn với 10 hapten nhỏ; nhưng đối với những kháng nguyên lớn, do sự hạn chế về không gian nên IgM chỉ có thể gắn với 5 phân tử trong cùng một thời điểm. Như vậy phân tử IgM có tính "hám" kháng nguyên cao hơn các loại globulin miễn dịch khác. Tính chất này của IgM đã tạo ra khả năng dễ kết hợp với các kháng nguyên đa chiều như các hạt virus và hồng cầu. Ví dụ khi hồng cầu được ủ với các kháng thể đặc hiệu chúng sẽ ngưng tập lại với nhau trong một quá trình được gọi là hiện tượng ngưng kết. Để xẩy ra cùng một mức ngưng kết thì lượng phân tử IgM cần thiết nhỏ hơn lượng phân tử IgG 100 đến 1.000 lần. Để trung hoà các virus thì lượng IgM cũng cần ít hơn lượng IgG. IgM có hiệu quả hơn IgG trong việc hoạt hoá bổ thể. Sự hoạt hoá bổ thể đòi hỏi phải có 2 mảnh Fc rất gần nhau, phân tử IgM đã đáp ứng được điều này vì vậy chúng hoạt hoá mạnh.
Do có kích thước lớn - trọng lượng phân tử 900.000, hằng số lắng 19S - nên IgM chỉ có trong lòng mạch và có nồng độ rất thấp ở dịch gian bào. Sự có mặt của chuỗi J làm cho phân tử có thể kết hợp với các thụ thể trên tế bào tiết và được chuyển vận qua hàng rào biểu mô vào dịch tiết. Mặc dù IgA là lớp kháng thể chính có trong dịch tiết, nhưng IgM cũng hoạt động như một globulin miễn dịch tiết bổ sung.
3.3. Kháng thể IgA
Mặc dù IgA chỉ chiếm 10 - 15% tổng lượng globulin miễn dịch trong huyết thanh, nhưng nó là lớp globulin miễn dịch chính trong dịch ngoại tiết như sữa, nước bọt, nước mắt, dịch nhầy khí phế quản, đường tiết niệu sinh dục, đường tiêu hoá. Trong huyết thanh IgA tồn tại dưới dạng monomer, nhưng đôi khi cũng tồn tại dưới dạng polymer như dimer, trimer và cả tetramer. IgA trong dịch ngoại tiết được gọi là IgA tiết, tồn tại dưới dạng dimer hoặc tetramer, có thêm chuỗi polypeptide J và một chuỗi polypeptide nữa được gọi là mảnh tiết.
Chuỗi J giống với chuỗi J của IgM pentamer cần thiết cho quá trình polymer hoá của IgA huyết thanh lẫn IgA tiết. Mảnh tiết là một polypeptide 70.000 Da do tế bào biểu mô của màng nhầy đường tiêu hoá, hô hấp, trong hốc mắt, tuyến nước bọt nhỏ, đường tiết niệu, và tử cung sản sinh ra. Lượng IgA tiết được sinh ra trong mỗi ngày lớn hơn lượng của bất kỳ globulin miễn dịch nào khác. Tế bào plasma tiết IgA tập trung ở bề mặt màng nhầy dọc theo hỗng tràng. Có khoảng 2,5x1010 tế bào plasma tiết IgA, lớn hơn số lượng tế bào plasma của tuỷ xương, hạch lympho và lách cộng lại. Mỗi ngày có khoảng 300 mg IgA tiết được tiết ra dọc theo hỗng tràng.
Mảnh tiết cần thiết cho sự chuyển vận IgA dimer qua tế bào biểu mô nhầy vào dịch tiết nhầy (hình 5.14). IgA có thể gắn một cách chặt chẽ với thụ thể trên bề mặt tế bào biểu mô nhầy dành cho phân tử globulin miễn dịch polymer. Phức hợp thụ thể - IgA sẽ được nhấn chìm về phía bào tương và nằm trong một bọng rồi được chuyển vận qua tế bào tới mặt phía trong lòng ống. Tại đây bọng sẽ liên hợp với màng plasma, sau đó thụ thể được phân cắt bởi enzyme và một phần thụ thể sẽ trở thành mảnh tiết. Mảnh tiết này được gắn vào và được giải phóng cùng với phân tử IgA dimer vào dịch tiết. Sự có mặt của mảnh tiết còn có tác dụng bảo vệ phân tử IgA không bị tác dụng của các enzyme thuỷ phân protein có trong dịch tiết phân huỷ. Chỉ có IgA dimer được chuyển vận qua tế bào biểu mô nhầy và người ta giả thiết rằng thụ thể dành cho globulin miễn dịch polymer đã nhận dạng chuỗi J của phân tử dimer. Vì vậy ngoài IgA tiết thì IgM pentamer cũng có chuỗi J nên cũng có thể được chuyển vận vào dịch tiết nhầy.
IgA tiết còn có một chức năng hết sức quan trọng trong việc sinh ra miễn dịch tại chỗ của đường tiêu hoá, đường hô hấp, tiết niệu sinh dục vì đây là những con đường chính để phần lớn các vi sinh vật gây bệnh xâm nhập vào cơ thể. IgA tiết gắn với các cấu trúc của bề mặt vi khuẩn hoặc virus và ngăn cản các vi sinh vật gắn vào tế bào nhầy. Vì vậy IgA tiết có tác dụng ngăn cản sự nhiễm virus và ức chế quá trình xâm nhập của vi khuẩn.
3.4. Kháng thể IgE
Mặc dù IgE có nồng độ trong huyết thanh rất nhỏ, chỉ 0,3 (g/ml nhưng người ta có thể nhận biết được qua hoạt động sinh học của chúng. Các kháng thể IgE gây ra các phản ứng quá mẫn thức thì với những tính chất của sốt rơm, hen, mầy đay, và sốc phản vệ. Năm 1921 Prausnitz .K và Kustner .H là những người đầu tiên chứng minh rằng có một thành phần trong huyết thanh gây ra các phản ứng dị ứng. Các tác giả đã lấy huyết thanh của một người bị dị ứng tiêm trong da cho một người không bị dị ứng, sau đó tiêm kháng nguyên thích hợp vào cùng vị trí tiêm huyết thanh thì thấy xuất hiện một quầng đỏ và sưng (giống như mầy đay). Phản ứng này được đặt tên là phản ứng PK (viết tắt của hai chữ Prausnitz và Kustner) đó là thử nghiệm sinh học đầu tiên để phát hiện hoạt tính của IgE. Năm 1966 hai vợ chồng nhà khoa học Ishizaka đã phát hiện một cách đích thực sự có mặt của IgE trong huyết thanh. Họ lấy huyết thanh từ một cơ thể bị dị ứng gây miễn dịch cho thỏ để thu được kháng huyết thanh kháng isotype. Kháng huyết thanh của thỏ được phản ứng với từng lớp kháng thể của người đã biết vào thời điểm đó, đó là IgG, IgA, IgM và IgD. Bằng cách này các kháng thể kháng isotype tương ứng với IgG, IgA, IgM và IgD sẽ bị tủa cùng với kháng nguyên tương ứng và được loại bỏ khỏi huyết thanh thỏ. Phần còn lại là kháng thể kháng isotype đặc hiệu với một lớp kháng thể chưa biết. Kháng thể kháng isotype này hoá ra lại phong bế hoàn toàn phản ứng PK. Kháng thể mới này được đặt tên là globulin miễn dịch E (chữ E là bắt nguồn từ kháng nguyên E của một loại phấn hoa có khả năng kích thích sinh ra lớp kháng thể này).
Phân tử IgE gồm hai chuỗi nặng epsilon và hai chuỗi nhẹ.Mỗi phân tử IgE có hai cầu disulfide nối chuỗi nặng với chuỗi nhẹ và hai cầu disulfide nối chuỗi nặng với chuỗi nặng. Trọng lượng phân tử 180.000, hằng số lắng 8S, thời gian bán phân huỷ khoảng 2 - 3 ngày. IgE rất dễ bị biến tính khi xử lý bằng các tác nhân khử hoặc bằng nhiệt. Ví dụ ở 56 ºC trong 30 phút thì IgE đã bị biến tính.
IgE gắn với các thụ thể dành cho Fc trên bề mặt bạch cầu ái kiềm ở máu ngoại vi và tế bào mast ở mô. Sau khi đã gắn lên bề mặt các tế bào này thì các vị trí kết hợp kháng nguyên ở phần Fab của IgE vẫn có thể gắn với kháng nguyên và kháng nguyên sẽ nối các phân tử IgE kề nhau lại. Sự liên kết chéo của các phân tử IgE đã gắn với thụ thể bởi kháng nguyên đã dẫn đến hiện tượng thoát bọng của bạch cầu ái kiềm và tế bào mast làm giải phóng các chất trung gian hoá học như serotonin, histamin. Các chất trung gian hoạt mạch này gây tăng tính thấm mao mạch giúp cho các kháng thể trong máu và các đại thực bào dễ dàng lọt qua thành mạch để đến những nơi mà kháng nguyên có thể xâm nhập vào cơ thể (da, niêm mạc). Do tác dụng làm tăng tính thấm mao mạch mà hệ thống [IgE - tế bào mast - các amin hoạt mạch] được ví như "người canh cửa" tại những nơi mà kháng nguyên có thể vào cơ thể. Tuy nhiên khi hiện tượng thoát bọng xẩy ra quá mạnh, lượng amine hoạt mạch được giải phóng quá nhiều và rầm rộ thì sẽ làm xuất hiện các triệu chứng dị ứng. Ngoài ra sự thoát bọng của tế bào mast bởi IgE cũng có thể làm giải phóng các chất trung gian hoá học có tác dụng chiêu mộ các loại tế bào khác nhau để chống lại ký sinh trùng.
3.5. Kháng thể IgD
IgD được phát hiện lần đầu tiên ở một bệnh nhân bị bệnh đa u tuỷ mà protein đa u tuỷ của bệnh nhân này không phản ứng với kháng huyết thanh kháng isotype kháng lại các isotype đã biết lúc đó là IgG, IgM và IgA. Nếu lấy protein đa u tuỷ này gây miễn dịch cho thỏ thì thu được kháng huyết thanh phản ứng với một lớp kháng thể mới có trong huyết thanh người bình thường với nồng độ thấp. Lớp kháng thể này được gọi là IgD có nồng độ khoảng 30 mg/ml huyết thanh chiếm 0,2% tổng lượng globulin miễn dịch huyết thanh.
Phân tử IgD có hai chuỗi nặng delta (d) và hai chuỗi nhẹ (hoặc k hoặc l) được nối với nhau bằng các cầu liên chuỗi, trong đó có hai cầu liên chuỗi nối chuỗi nặng với chuỗi nhẹ và một cầu nối chuỗi nặng với chuỗi nặng. Trọng lượng phân tử 170.000 - 200.000, hằng số lắng là 7S. Tốc độ tổng hợp kém hơn IgG 100 lần, nhưng dị hoá nhanh (thời gian bán phân huỷ là vài ba ngày) và rất dễ bị thuỷ phân bởi enzyme plasmin trong quá trình đông máu. IgD cũng rất dễ bị biến chất bởi nhiệt và acid ngay cả ở mức độ mà IgG, IgA hoặc IgM không bị ảnh hưởng gì.
Cho đến nay người ta chưa rõ chức năng sinh học của IgD. Trong huyết thanh những người bị nhiễm khuẩn mạn tính có IgD tăng, nhưng không đặc hiệu cho một loại nào. IgD có trong kháng thể kháng nhân, kháng tuyến giáp, kháng insulin, kháng penicilin, kháng độc tố bạch cầu. IgD không kết hợp bổ thể, không gây phản vệ thụ động trên da chuột lang và không qua được nhau thai. Điều đáng chú ý là IgD cũng là một lớp Ig xuất hiện trên màng các tế bào B chín và vì vậy người ta nghĩ rằng nó có chức năng trong quá trình hoạt hoá tế bào B bởi kháng nguyên.
Receive articles via Email!