Tìm Hiểu Về Sự Động Dục Trên Chó Cái

1.1. CƠ QUAN SINH DỤC CỦA CHÓ CÁI
1.1.1. Chức năng
Cơ quan sinh dục cái đảm nhận các chức năng sinh học:
- Sản xuất noãn bào (trứng)
- Vận chuyển noãn bào từ buồng trứng theo đường ống dẫn trứng đến vị trí thụ tinh
- Dự trữ và hoàn thiện khả năng thụ tinh của tinh trùng
- Định vị và nuôi dưỡng phôi thai
- Sinh con
- Tổng hợp và phân tiết kích thích tố sinh dục cái estrogen và progesterone
1.1.2. Cấu tạo
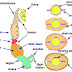
Cơ quan sinh dục chó cái gồm dây rộng, buồng trứng, ống dẫn trứng, tử cung, âm đạo và âm hộ.
1.1.2.1. Dây rộng
Dây rộng là những nếp gấp phúc mô treo các cấu tạo của cơ quan sinh dục cái bên trong, ngoại trừ âm đạo. Mỗi dây rộng chia làm 3 phần:
- Màng treo buồng trứng là phần dây rộng tạo nên vách phía trong của túi buồng trứng. Túi buồng trứng là túi phúc mô mỏng, bao bọc buồng trứng, thông vào xoang phúc mô qua một khe hẹp.
- Màng treo ống dẫn trứng là phần nối tiếp với màng treo buồng trứng, bám vào ống dẫn trứng và cùng với màng treo buồng trứng tạo thành túi buồng trứng.
- Màng treo tử cung bắt nguồn từ phần xương chậu và vùng thắt lưng để bám vào cạnh của đoạn trước âm đạo, cổ tử cung, thân và sừng tử cung tương ứng.
1.1.2.2. Buồng trứng
Buồng trứng của chó cái gồm một đôi, có hình ovan đến hình tròn, nằm trong hai túi buồng trứng, ở phía sau thận. Mỗi buồng trứng được đính bởi dây riêng vào tử cung và dây treo vào cân mạc ngang cửa bụng, ngay phía trong của xương sườn chót, khoảng đốt sống thắt lưng thứ 3 hoặc thứ 4. Buồng trứng phải thường nằm về trước hơn buồng trứng trái (vị trí khoảng 1/3 dưới thận trái). Buồng trứng vừa là tuyến ngoại tiết sản xuất tế bào sinh dục cái (noãn bào), vừa là tuyến nội tiết tổng hợp và phân tiết kích thích tố estrogen, progesterone.
1.1.2.3. Ống dẫn trứng
Ống dẫn trứng còn gọi là ống tử cung hay ống Fallope, bao bọc bởi túi buồng trứng. Phần đầu ống tiếp giáp với buồng trứng có dạng hình phễu được gọi là vòi Fallope và tận cùng ở phần tiếp giáp với sừng tử cung. Cửa ngõ đi vào sừng tử cung gọi là vòi tử cung.
1.1.2.4. Tử cung
Tử cung của chó cái có dạng chữ Y; gồm hai sừng tử cung, thân tử cung và cổ tử cung. Tử cung định vị ở khoảng giữa phần bụng của bàng quang và kết tràng xuống (một phần nằm trong xoang bụng và một phần trong xoang chậu). Kích thước của tử cung rất thay đổi, phụ thuộc vào tầm vóc của thú, số lần mang thai, tình trạng bệnh lý sinh sản, chó cái có mang thai hay không mang thai.
Sừng tử cung là một ống màng cơ hơi hẹp từ vùng lưng xuống bụng, tiếp nối với ống dẫn trứng ở phía trước và thân tử cung ở phía sau. Sừng tử cung nằm hoàn toàn trong xoang bụng, sừng bên phải thường dài hơn sừng bên trái. Thân tử cung nằm trong xoang bụng và một phần trong xoang chậu, phía trước tiếp nối với 2 nhánh của sừng tử cung và phía sau là âm đạo thông qua cổ tử cung. Cổ tử cung là phần thu hẹp của thân tử cung tiếp nối với âm đạo.
1.1.2.5. Âm đạo
Âm đạo nằm giữa cổ tử cung và tiền đình, hoàn toàn trong xoang chậu. Phần đầu âm đạo được gọi là vòm âm đạo, phần còn lại kéo dài về phía trước có lớp nội bì xếp theo chiều dọc và các nếp gấp nhỏ xếp theo chiều ngang. Nếp dọc tận cùng ở ngang tầm với lỗ thoát tiểu, là nơi tiếp nối với tiền đình. Âm đạo đảm nhận các chức năng như tiếp nhận dương vật của thú đực trong quá trình phối giống và là đường tiếp dẫn thú con sinh ra.
1.1.2.6. Âm hộ
Âm hộ là cửa ngỏ của cơ quan sinh dục cái, gồm 2 môi, một ống niệu dục ngoài và một khe thẹn (âm môn).
1.2. SỰ PHÁT TRIỂN NANG NOÃN VÀ NOÃN BÀO
1.2.1. Sự phát triển nang noãn
Dưới kính hiển vi, miền vỏ buồng trứng có nhiều nang noãn ở các giai đoạn phát triển khác nhau, bao gồm các dạng nang noãn:
- Nang noãn nguyên thủy: Ở buồng trứng thú non có nhiều nang noãn nguyên thủy, nhưng phần lớn không phát triển và dần dần bị tiêu hủy. Chúng xếp thành từng đám như ở buồng trứng của chó và mèo hoặc rải rác như ở buồng trứng thú nhai lại. Cấu tạo của mỗi nang noãn nguyên thủy gồm noãn bào bậc I bao bọc bởi lớp tế bào nang dẹp, ngoài là lớp màng đáy mỏng. Tế bào nang có nguồn gốc từ tế bào sợi của mô liên kết, với nhiệm vụ của tế bào nang là cung cấp dưỡng chất giúp cho noãn bào phát triển.
- Nang noãn sơ cấp: Các nang noãn sơ cấp có kích thước to nhỏ khác nhau và bên trong m, tiến trình tổng hợp chấtmlà noãn bào bậc I đang gia tăng số lượng tế bào, kích thước noãn bào có thể đạt 125-150 trong noãn bào xảy ra với cường độ cao. Giữa noãn bào và lớp tế bào hạt xuất hiện một lớp màng trong suốt được gọi là màng thấu quang, ngoài là lớp màng đáy.
màng xếp thành nhiều lớp tế bào, m Các tế bào hạt vào giai đoạn này có dạng hình khối đơn hay đa diện với kích thước khoảng 8 - 10 trong tế bào chất của tế bào nang chứa nhiều ty thể, bộ Golgi, lưới võng nội bào có hạt và các hạt lipid. Trong quá trình phát triển của nang noãn sơ cấp, một số tế bào nang gia tăng số lượng bằng hiện tượng phân bào đẳng nhiễm (mitosis) hình thành nhiều lớp bao quanh noãn bào bậc I và các tế bào này có khả năng tổng hợp và phân tiết estrogen để kích thích sự phát triển và thành thục của noãn bào. Ở mỗi chu kỳ, chỉ có một ít nang noãn sơ cấp phát triển tiếp tục tạo thành nang noãn thứ cấp.
- Nang noãn thứ cấp: Những nang noãn này càng tăng kích thước với lớp tế bào nang hình khối hoặc hình trụ phát triển mạnh, có nhiều hạt trong tế bào chất, tạo nhiều lớp gọi là lớp tế bào hạt ngăn cách với noãn bào bằng màng thấu quang và chúng bắt đầu phân tiết dịch nang.
Giai đoạn này được đánh dấu bởi các tế bào bản thể phát triển tạo thành 2 lớp vỏ bao bọc nang noãn. Lớp vỏtrong được cấu tạo bởi các tế bào có kích thước lớn, tròn và giống như các tế bào biểu mô, chúng có khả năng tổng hợp và phân tiết các hormone androgen và được chuyển hoá thành estrogen bởi các tế bào hạt. Các tế bào của lớp vỏ ngoài thì nhỏ hơn, cấu tạo như sợi nguyên bào.
- Nang noãn trưởng thành hay nang Graaf: Nang noãn thứ cấp tiếp tục gia tăng kích thước, lớp tế bào hạt phát triển và xuất hiện các khe chứa dịch nang. Chất dịch càng nhiều thì các khe càng nở to và hợp thành một hốc lớn duy nhất gọi là xoang nang.
Hốc nang được bao quanh bởi lớp tế bào hạt áp sát noãn bào từ dạng khối đơn dần dần chuyển sang trụ đơn, xếp thành vùng phóng xạ, có vai trò cung cấp dưỡng chất cho noãn bào (noãn bào bậc I). Noãn bào được bao bọc bởi màng thấu quang, vùng phóng xạ và một số tế bào hạt họp thành đĩa trứng. Bao quanh màng hạt là lớp mô liên kết có chứa nhiều mao mạch gọi là màng nang.
- Nang noãn thành thục: Những nang Graaf có kích thước to lớn, trồi lên bề mặt buồng trứng. Bên trong, m và có khảmnoãn bào đã phát triển rất lớn, có thể đạt kích thước 200 năng thụ tinh. Tuy nhiên, đối với chó, nang noãn thành thục có chứa noãn bào bậc I chưa có khả năng thụ tinh.
Bình thường ở thú đơn thai, trong một chu kỳ động dục chỉ có khoảng 12 nang noãn thành thục trồi lên bề mặt buồng trứng. Riêng ở chó, mèo, heo, cừu thì số lượng nang noãn thành thục có thể nhiều hơn. Sau khi xuất noãn, vết nứt trên bề mặt buồng trứng chứa đầy máu trong xoang gọi là hồng thể. Cấu tạo này phát triển và tích lũy sắc tố màu vàng tan trong lipid, chúng được gọi là hoàng thể, dưới tác động của LH kích thích các tế bào hạt tổng hợp và phân tiết progesterone. Hoàng thể sẽ tồn tại trong suốt giai đoạn thú mang thai, nếu thú không mang thai thì sẽ thoái hoá sau 30 ngày và tạo vết sẹo gọi là bạch thể
Sự biến thiên hàm lượng kích thích tố buồng trứng trong huyết thanh và tác động sinh học qua các giai đoạntrong chu kỳ động dục được trình bày ở phần 1.4.3 và 1.4.4.
1.2.2. Sự phát triển noãn bào
Vào giai đoạn phôi, các tế bào mầm nguyên thủy di chuyển từ túi noãn hoàng đến tập trung tại chồi sinh dục (phát triển tạo thành buồng trứng, tại đây các tế bào mầm phát triển tạo thành noãn nguyên bào 2n)
Noãn nguyên bào phân bào đẳng nhiễm và sau khi hoàn thành lần phân bào đẳng nhiễm đầu tiên tạo thành noãn bào bậc I (2n) thì chúng không tiếp tục phân chia mà chỉ phát triển, tích lũy dưỡng chất và gia tăng kích thước; giai đoạn này kéo dài từ giai đoạn thai đến khi thú trưởng thành sinh dục. Vào giai đoạn thành thục tính dục, dưới tác động của FSH, một số lượng nhỏ noãn bào bậc I (2n) được kích thích phân bào theo kiểu gián phân giảm nhiễm I (meiosis I), tạo thành noãn bào bậc II (n) và thể cực (n). Thể cực (n) là một tế bào có nhân, có n nhiễm sắc thể, rất ít tế bào chất và sau đó chúng bị thoái hoá.Trong hầu hết các loài động vật, giai đoạn phân bào giảm nhiễm hoàn thành (meiosis II) trước khi có hiện tượng xuất noãn (Lâm thị Thu Hương, 2005 [3]).
Theo Feldman , ở chó, noãn bào khi rụng là noãn bào bậc I chưa hoàn toàn thành thục và không có khả năng thụ tinh; chúng sẽ hoàn toàn thành thục tạo thành noãn bào bậc II (n) và có khả năng thụ tinh trong khoảng thời gian ít nhất là 48 giờ trong ống dẫn trứng.
1.3. CHU KỲ SINH DỤC CỦA CHÓ CÁI
1.3.1. Sự thành thục tính dục
Tuổi thành thục hay trưởng thành sinh dục là tuổi mà cơ quan sinh dục bắt đầu hoạt động, buồng trứng của thú cái sản sinh noãn bào, dịch hoàn sản sinh tinh trùng và các tế bào giao tử này có khả năng thụ tinh. Chó cái thành thục tính dục trung bình 6 - 12 tháng tuổi nhưng rất biến động và chịu tác động của nhiều yếu tố, trong đó giống có ảnh hưởng rất lớn đến thời điểm động dục lần đầu. Do vậy rất khó xác định chính xác tuổi thành thục của chó cái.
Concannon (1991) đã ghi nhận kinh nghiệm của các nhà chăn nuôi và cho biết các giống chó tầm vóc nhỏ thành thục khoảng 6 - 10 tháng tuổi, các giống tầm vóc to thành thục muộn hơn (khoảng 12 tháng tuổi); tuy nhiên, nhiều giống chó to có chu kỳ động dục đầu tiên xảy ra từ 18 - 24 tháng tuổi. Sự thay đổi tự nhiên này có thể do chó cái biểu hiện động dục thầm lặng nên nhà chuyên môn và chủ nuôi không thể xác định thời điểm động dục đầu tiên. Tương tự, Brooks (2003) , Hopkins (2003) ; Schulman (2004) nhận định các giống chó nhỏ thành thục tính dục rất sớm nhưng biến động và chó cái chỉ biểu hiện tính dục sau 2 - 3 tháng khi đạt thể vóc trưởng thành. Ngoài ra, trọng lượng cơ thể ảnh hưởng đến tuổi thành thục, chó nặng dưới 15kg thành thục khoảng 6 - 8 tháng tuổi, trong khi các chó nặng hơn sẽ động dục khoảng 8 - 10 tháng tuổi và khả năng thụ tinh thành công thường rơi vào chu kỳ thứ hai hoặc thứ ba. Từ kết quả khảo sát một số chỉ tiêu sinh lý sinh sản trên 50 chó cái German Shepherd, Nguyễn Văn Thanh (2005) ghi nhận 60% chó cái thành thục về tính tập trung từ 11- 13 tháng tuổi.
1.3.2. Các giai đoạn trong chu kỳ động dục
Từ khi bắt đầu trưởng thành tính dục, thú cái có những biểu hiện lên giống và quá trình biểu hiện này được lập đi lập lại có tính chu kỳ, khoảng cách giữa lần lên giống này với lần lên giống kế tiếp được gọi là chu kỳ động dục.
Chu kỳ động dục của chó cái phân chia 4 giai đoạn: trước động dục, động dục, sau động dục và nghỉ ngơi. Giai đoạn nghỉ ngơi và trước động dục thuộc pha nang noãn hay pha estrogen, giai đoạn động dục và sau động dục thuộc pha hoàng thể hay pha progesterone. Thời gian của chu kỳ và của mỗi giai đoạntrong chu kỳ động dục của chó rất biến động, thay đổi theo giống, điều kiện chăm sóc, quản lý, nuôi dưỡng và bệnh lý... (Johnston, 2000 ; Davol, 2000 ; Eilts, 2001 ; Correa, 2002).
1.3.2.1. Giai đoạn trước động dục
Giai đoạn trước động dục được xác định bắt đầu từ thời điểm có dấu hiệu sưng phồng và dịch tiết ở âm hộ đến khi chó cái cho phép chó đực nhảy chồm và phối giống. Những tiêu chuẩn có thể được sử dụng để xác định giai đoạn trước động dục của chó cái như âm hộ sưng lớn, dẫn dụ và thay đổi hành vi trước chó đực; ngoài ra, âm hộ tiết nhiều dịch nhầy có lẫn máu. Xét nghiệm tế bào âm đạo ghi nhận sự thay đổi hình dạng tế bào biểu mô và sự hiện hữu của các loại tế bào khác như hồng cầu, bạch cầu... Có thể xác định chắc chắn thời điểm bắt đầu của giai đoạn trước động dục là khi nhìn thấy âm hộ chó cái sưng phồng và tiết dịch có lẫn máu. Giai đoạn trước động dục kéo dài 6 đến 11 ngày, trung bình 9 ngày; tuy nhiên, khoảng thời gian này rất biến động, có thể 2 ngày hoặc 25 ngày (Nelson, 1998 ).
Vào đầu giai đoạn trước động dục, chó cái có những hành vi như đùa nghịch, trêu chọc các chó khác, nhưng không cho phép chó đực tiếp xúc hay chồm nhảy với biểu hiện gầm gừ, tránh né liên tục, đuôi cụp lại giữa hai chân sau hoặc nằm bẹp xuống. Sang cuối giai đoạn trước động dục, hành vi của chó cái trở nên thụ động hơn, có thể đứng hoặc ngồi khi chó đực ve vãn. Trong giai đoạn này, hiện tượng xuất dịch nhầy lẫn máu rất thay đổi theo cá thể, có cá thể xuất huyết nhiều và có cá thể xuất huyết ít. Máu hiện diện trong dịch thải là do sự thoát mạch của hồng cầu từ hệ thống mao quản của lớp nội mạc tử cung hay lớp dưới biểu mô, chảy qua cổ tử cung vào trong âm đạo. Kích thước của âm hộ lớn dần trong giai đoạn trước động dục là do hiện tượng sưng phồng ở hai môi âm hộ. Sự sưng phồng âm hộ của chó cái đã gây trở ngại cho việc chó đực đưa dương vật vào. Khi bước sang giai đoạn động dục thì âm hộ giảm sưng, mềm hơn, dịch thải trong và keo lại (Feldman và Nelson, 1996 ; Correa, 2002 ).
1.3.2.2. Giai đoạn động dục
Davol (2000) ; Hopkins (2003) xác nhận thời gian động dục của chó cái rất biến động, thay đổi theo từng cá thể, từ 3 - 21 ngày và trung bình 9 ngày, sự xuất noãn sẽ xảy ra sau 48 giờ khi bước qua giai đoạn động dục.
Theo Feldman và Nelson (1996) , thời gian động dục trung bình ở chó cái từ 5 - 9 ngày, tuy nhiên có cá thể động dục kéo dài khoảng 1- 2 ngày, nhưng có những cá thể lại động dục kéo dài đến 18 - 20 ngày. Vì thế, rất khó dự đoán chính xác thời gian động dục của chó.
Hành vi của chó cái thay đổi ở giai đoạn động dục bao gồm tiếp xúc và cho phép chó đực nhảy chồm để chuẩn bị phối giống. Các chó cái này biểu hiện cúi mình và đưa âm hộ ra phía trước chó đực. Thông thường chó cái ở vào giai đoạn này phân tiết nhiều pheromone nên có thể hấp dẫn chó đực dù ở khoảng cách xa, vì thế hình ảnh thường gặp khi chó cái động dục là nhiều chó đực ve vãn một chó cái (Schulman, 2004 ).
Âm hộ bớt sưng phồng, mềm và dịch thải thường có màu dâu hoặc màu hồng, rất ít khi thấy lẫn máu. Trong dịch thải có chứa glucose và có thể xác nhận bằng cách xét nghiệm que thử nước tiểu. Glucose hiện diện trong dịch thải là do tác động của progesterone kháng insulin nên làm tăng hàm lượng đường huyết, đồng thời nâng cao lượng glucose trong các dịch ngoại bào (Feldman và Nelson, 1996 ).
Nguyễn Văn Thanh (2005) theo dõi trên 20 chó cái German Shepherd ở giai đoạn động dục và ghi nhận thân nhiệt trung bình 39,63 - 39,850C; tương ứng vào ngày thứ 12 - 14, là thời điểm mà số lượng noãn bào chín và xuất cao nhất, nếu phối giống vào thời điểm này cho tỉ lệ thụ tinh cao (80%).
1.3.2.3. Giai đoạn sau động dục
Giai đoạn sau động dục thuộc pha progesterone do hàm lượng progesterone huyết thanh gia tăng trong khoảng thời gian này và kéo dài khoảng 56 - 58 ngàytrong lúc mang thai (Olson và ctv., 1984 ).
Correa (2002); Schulman (2004) xác định giai đoạn sau động dục của chó kéo dài 2 tháng, từ khi chó cái không biểu hiện hấp dẫn chó đực và chấm dứt khi hoàng thể thoái hoá hoàn toàn.
Dấu hiệu rõ nhất được ghi nhận ở giai đoạn sau động dục là sự từ chối phối giống, không lôi cuốn chó đực. Âm hộ trở về trạng thái và kích thước bình thường, không mềm nhũn như ở giai đoạn động dục. Thực tế cho thấy không thể dựa vào biểu hiện lâm sàng để phân định chó ở giai đoạn sau động dục hay nghỉ ngơi hoặc những ngày đầu thai kỳ (Feldman và Nelson, 1996 ; Davol, 2000 ).
1.3.2.4. Giai đoạn nghỉ ngơi
Tương tự như các giai đoạn khác trong chu kỳ động dục của chó cái, thời gian của giai đoạn này rất biến động; tùy thuộc vào giống, tình trạng sức khoẻ, thời gian trong năm, môi trường và các yếu tố phức tạp khác. Một chu kỳ động dục tiêu biểu của chó cái khoảng 7 tháng, bắt đầu giai đoạn trước động dục kéo dài 9 ngày, động dục 7 - 9 ngày, sau động dục 58 ngày và nghỉ ngơi 4,5 tháng (Davol, 2000 ).
Nhiều tác giả đã cho rằng dựa vào hàm lượng progesterone huyết thanh có thể là căn cứ chính để xác định giao điểm giữa hai chu kỳ động dục kế tiếp trên các chó cái không mang thai.
Để rút ngắn thời gian của pha hoàng thể hay thời gian sau động dục từ vài tuần đến vài tháng có thể sử dụng prostaglandin (PGF2α) gây phân giải hoàng thể hoặc cung cấp bromocriptine một thời gian dài (Hess, 2002 ; Phillips và ctv., 2003).
Klein và ctv (2003) cung cấp estradiol benzoate trên 5 chó cái Beagle (1- 3 năm tuổi) vào giai đoạn nghỉ ngơi với mục đích chấm dứt giai đoạn nghỉ ngơi và thúc đẩy quá trình động dục kế tiếp. Tác giả khảo sát hàm lượng FSH, LH và estrogen huyết thanh trong 7 ngày liên tiếp và ghi nhận estrogen có tác động hồi phản dương đến vùng dưới đồi. Kết quả cho thấy nhóm chó thí nghiệm có hàm lượng LH huyết thanh gia tăng vào thời điểm trước khi xuất noãn và chấm dứt giai đoạn nghỉ ngơi của chu kỳ trước.
 |
| Cơ chế kiểm soát quá trình tổng hợp và phân tiết kích thích tố sinh sản. |
Ngoài ra, do hậu quả của quá trình mang thai và sinh con nên khoảng thời gian nghỉ ngơi giữa hai chu kỳ động dục hiếm khi bất biến.
Hành vi và dấu hiệu sinh lý của chó cái ở giai đoạn nghỉ ngơi hoàn toàn bình thường, không đặc trưng; đồng thời, không thể phân biệt lâm sàng giữa chó cái ở giai đoạn nghỉ ngơi với chó ở cuối giai đoạn sau động dục hoặc cắt bỏ buồng trứng. Đối với chó bị cắt bỏ buồng trứng, hàm lượng FSH và LH huyết thanh tăng cao hơn so với chó ở giai đoạn không động dục (Feldman và Nelson, 1996 ).
1.4. KÍCH THÍCH TỐ BUỒNG TRỨNG KIỂM SOÁT CHU KỲ SINH DỤC
1.4.1. Sự tổng hợp và phân tiết kích thích tố buồng trứng
Sự tổng hợp và phân tiết kích thích tố estrogen và progesterone chịu sự kiểm soát của kích thích tố sinh dục não thùy (FSH - Follicle stimulating hormone và LH - Luteinizing hormone) dưới tác động của GnRH (Gonadotropin - releasing hormone) phân tiết từ vùng dưới đồi (Messer, 2000 ; Kimball, 2002 ).
Kích thích tố sinh dục não thuỳ (gonadotropin hormone) được tổng hợp và phân tiết từng nhịp dưới sự kích thích của GnRH từ vùng dưới đồi đã được Belchetz và ctv. (1978) nghiên cứu ở khỉ, Clarke và Cummins (1982) nghiên cứu ở cừu, Irvine và Alexander (1986) khảo sát ở ngựa (trích dẫn của Mol và Rijnberk, 1997 ).
Trong công trình nghiên cứu chức năng của hoàng thể trong suốt giai đoạn sau động dục, Fernandes và ctv. (1987) nhận định hàm lượng LH và prolactin huyết thanh có tương quan thuận với số lượng thụ thể LH và thụ thể prolactin ở hoàng thể.
Tuy nhiên, Padmanabhan và ctv. (1997) cho rằng rất khó xác định mối tương quan giữa GnRH với cung lượng FSH bởi vì thời gian phân tiết FSH kéo dài vào giai đoạn nghỉ ngơi, thú cái không động dục và FSH có thời gian bán rã trung bình dài hơn LH (Eilts, 2001 ). Tác giả ghi nhận FSH được tổng hợp và phân tiết từng nhịp từ não thùy trước ở cừu cái bị cắt bỏ buồng trứng khi cung cấp GnRH.
Ở động vật cái, FSH có vai trò kích thích sự phát triển và chín của các nang noãn trong buồng trứng. LH có tác động kích thích sự sản sinh androstenedione ở lớp vỏ trong, chất này được chuyển thành 17β-estradiol; sau đó, estradiol tác kích từng đợt lên vùng dưới đồi, thúc đẩy sự phân tiết GnRH. Khi GnRH đạt đỉnh, não thùy trước phân tiết LH cao độ, mở đầu tiến trình xuất noãn. Vizcarra và ctv. (1997) ghi nhận vấn đề này qua khảo sát 32 bò cái được cung cấp GnRH, tác giả xem xét ảnh hưởng của GnRH đến sự phát triển nang noãn, hàm lượng LH và FSH huyết thanh, số lượng thụ thể LH, FSH, GnRH và mRNA mã hoá GnRH ở tuyến não thùy trước.
 |
| Sự biến thiên hàm lượng kích thích tố sinh dục và tế bào trong chu kỳ động dục |
Kooistra và ctv. (1999) nghiên cứu sự phân tiết FSH và LH trên đàn chó Beagle và ghi nhận hàm lượng hai loại kích thích tố này biến động từng hồi, từng pha khác nhautrong giai đoạn động dục và giai đoạn nghỉ ngơi.
Một khảo sát của Margaret và ctv. (2004) về sự thay đổi hàm lượng gonadotropin trên 10 chó cái Beagle bị cắt bỏ buồng trứng. Hàm lượng các gonadotropin huyết tương gia tăng nhanh chóng trong 3 - 5 tuần sau khi cắt bỏ buồng trứng, nhưng sụt giảm đột ngột vào tuần 10 và tăng vào tuần 42. Hàm lượng FSH và LH huyết tương trung bình của các chó cái thí nghiệm sau 12 tháng lần lượt là 75,3 ng/ml và 8,3 ng/ml.
Margaret và ctv. (2005) so sánh hàm lượng FSH và LH huyết tương trên nhóm chó cái bình thường và nhóm chó cái bị thiến ở giai đoạn nghỉ ngơi. Hàm lượng FSH và LH huyết tương ở nhóm chó thiến tăng gấp 10 lần so với nhóm chó bình thường (FSH 62,5 ng/ml so với 5,2 ng/ml và LH 6,1 ng/ml so với 0,5 ng/ml).
Ngoài ra, sự phân tiết progesterone từ hoàng thể chịu sự kiểm soát của một số kích thích tố khác như GH (Growth hormone), PRL (Prolactin), prostaglandin và các chất β-adrenergic. Hơn nữa, follistatin từ các tế bào hạt có khả năng liên kết với activin để điều hoà phân tiết progesterone
1.4.2. Cấu tạo và chức năng sinh học của các kích thích tố buồng trứng
Kích thích tố buồng trứng thuộc nhóm steroid, gồm estrogen (18 carbon) và progesterone (21 carbon) (Edqvist và Forsberg, 1997 ; Bulun và ctv., 2000 ; Frank, 2003 ).
1.4.2.1. Estrogen
Estrogen vận chuyển trong máu dưới dạng kết hợp với globulin huyết thanh (SHBG - sex hormone binding globulin) (Kahn và ctv., 2002 ) và thể hiện tác động sinh học trong nhân của tế bào đích thông qua sự liên kết với thụ thể estrogen đặc hiệu (ER ). Bộbvà ER-a- estrogen receptor). Thụ thể estrogen gồm hai loại (ER- ở nhiễm sắc thể thứ 1bđịnh vị ở nhiễm sắc thể thứ 6 và ER-agen ER-4 của người.
Theo dẫn liệu Molenda và ctv. (2003) ; Ververidis và ctv. (2004) , thụ thể estrogen có vai trò quan trọng đối với tác động sinh học của estrogen. Chúng hiện diện ở nhiều loại mô, từ hệ thống thần kinh trung ương, hệ thống tuần hoàn, miễn dịch, hô hấp (phổi), tiêu hoá (dạ dày và ruột), tiết niệu (thận và đường niệu dục), đến mô xương. Đối với hệ thống thần kinh trung ương, estrogen có tác động kiểm soát quá trình sinh lý sinh sản của động vật cái bao gồm hành vi, biểu hiện được tìm thấy ở trung khu khứu giác và ởbtrạng thái sinh lý, bởi vì ER- vùng vỏ tiểu não.
Horard và Vanacker (2003) đề cập đến các thụ thể liên quan của ER (ERR-estrogen receptor-related receptors). ERR được gọi là các thụ thể “cô nhi” (orphan receptors), chúng hiện diện trong và ERR-γ,b, ERR-anhân tế bào, gồm ba loại là ERR- trong đó kiểm soát sự biểu hiện gen trong quá trình đáp ứng với các tín hiệu sinh học từ estrogen.
De Brito và ctv. (2006) ứng dụng kỹ thuật miễn dịch hoá mô xác định tính cảm ứng giữa 17-β estradiol và progesterone đối với thụ thể estrogen ở âm đạo và ở mô bướu truyền lây do giao phối (TVT - Transmissible venereal tumor). Kết quả cho thấy ER-α trên các chó cái bình thường ở giai đoạn nghỉ ngơi, trước động dục và động dục cảm ứng cao hơn giai đoạn sau động dục. Trong khi ở nhóm chó cái bị bướu, ER-α không biểu hiện cảm ứng ở mô bướu.
Theo Britt và Findlay, 2002 , tác động của estrogen trong sinh sản của động vật có vú gồm 2 tiến trình; đó là tạo nang noãn và duy trì kiểu hình của các tế bào bản thể ở buồng trứng.
(1). Tiến trình tạo nang noãn
Estrogen có mối liên quan mật thiết với FSH trong tiến trình sinh nang noãn và FSH là chìa khoá của tiến trình này. FSH phối hợp cùng yếu tố tăng trưởng giống insulin-1 (IGF-1, insulin-like growth factor-1) và estradiol (E2) để kích thích các tế bào hạt phát triển và biệt hoá. FSH kích thích phản ứng phosphoryl hoá trên cấu tử serine của protein kinase, giúp cho protein kinase hoạt động, từ đó kích động đến hệ thống IGF-1/IP3-K (insulin-like growth factor-1/inositol triphosphate - kinase) phụ thuộc vào PDK-1 (phosphatidyl inositol - dependent kinase 1) để thúc đẩy quá trình sao chép gen, tăng cường tổng hợp các men biến dưỡng trong tế bào hạt. Tuy nhiên, chuỗi liên hợp tác động trên chỉ được thể hiện khi E2 liên kết với ER-β trên màng tế bào hạt.
Thực nghiệm chứng minh nhóm chuột ER-α knock out (ERKO), ER-β knock out (BERKO) và ER-αβ knock out không có khả năng đáp ứng với E2 nội sinh hoặc ngoại sinh, trong khi nhóm aromatase knock out (ArKO) lại có khả năng đáp ứng với E2 ngoại sinh nhưng không chịu ảnh hưởng của E2 nội sinh. Nhóm chuột này đã có biểu hiện buồng trứng triển dưỡng, không hình thành hoàng thể, tiến trình sinh nang noãn bị chận đứng và các nang noãn thứ cấp biến đổi thành các nang tích máu
(2). Kiểu hình của các tế bào bản thể ở buồng trứng
Theo Feldman và Nelson, 1996 ; Meyers-Walen (2001, 2004) , từ lâu người ta đã biết rằng phái tính của loài động vật có vú được quyết định bởi cặp nhiễm sắc thể XX hoặc XY, sự liên quan này xảy ra vào giai đoạn phôi thai và bị chi phối bởi nhiều yếu tố sinh học của môi trường như nhiệt độ và nhất là do tác động của men aromatase.
Meyers-Wallen (2001) ; Smith (2001) ; Buergelt (2004) ; Domit (2004) ghi nhận chó có tổng số 78 nhiễm sắc thể, trong đó 38 cặp không liên quan đến phái tính và 2 nhiễm sắc thể phái tính; ở con cái, nhiễm sắc thể giới tính là XX và con đực là XY. Phái tính của động vật có vú được xác định vào thời điểm thụ tinh, dựa vào nhiễm sắc thể giới tính X và Y trong tinh trùng. Phôi thai có phái tính đực nếu tinh trùng thụ tinh có nhiễm sắc thể Y và ngược lại; đồng thời, cơ quan sinh dục của thú con được hình thành rất sớm từ giai đoạn phôi thai. Đối với phôi thai chỉ có nhiễm sắc thể X, buồng trứng phát triển từ tuyến sinh dục chưa phân hoá, cùng với hệ thống ống müllerian như vòi Fallop, tử cung, âm đạo và âm hộ. Đối với phôi thai có nhiễm sắc thể Y, tuyến sinh dục chưa phân hoá sẽ phát triển thành dịch hoàn, dịch hoàn sản sinh testosterone và MIS (müllerian inhibiting substance). MIS phân tiết từ các tế bào Sertoli và có tác động gây thoái hoá hệ thống ống dẫn müllerian. Testosterone được tổng hợp và phân tiết từ gian bào Leydig, chúng có tác động kích thích sự thành lập lớp biểu mô mầm và các tuyến sinh dục phụ từ hệ thống ống wolffian như ống dẫn tinh, nhiếp hộ tuyến, dương vật, bìu dái...
Vai trò của estrogen từ tuyến sinh dục trong sự biệt hoá và duy trì kiểu hình của các tế bào bản thể buồng trứng ở loài động vật có vú chưa xác định rõ. Tuy nhiên, thực nghiệm trên chuột thuộc nhóm ER-αβ knock out và ghi nhận các cấu trúc giống như các ống sinh tinh xuất hiện trong buồng trứng. Điều đó chứng tỏ estrogen có vai trò quan trọng đối với kiểu hình của các tế bào bản thể ở buồng trứng và quyết định đến phái tính của động vật (Britt và Findlay, 2002).
Theo Meyers-Wallen và Patterson (1989) (trích dẫn của Feldman và Nelson, 1996 ) biểu thị tình trạng chuyển đổi giới tính ở chó gồm nhiều trường hợp:
Kiểu gen giới tính (genotypic sex) hay kiểu nhiễm sắc thể giới tính (chromosomal sex type) phản ảnh nhiễm sắc thể giới tính, với các trường hợp XX, XY, XXY, XO...
Kiểu hình giới tính (phenotypic sex) dựa vào biểu hiện ở bộ phận sinh dục ngoài, chó cái có âm hộ và chó đực có dương vật. Tuy nhiên, kiểu hình giới tính chỉ có thể kết luận chính xác cần xác định: vị trí và hình dạng của âm hộ hay bao qui đầu,trong âm hộ có âm vật hay trong bao qui đầu có dương vật hay không, vị trí của cơ quan sinh dục khi mở ra, và chó có nhiếp hộ tuyến hay âm đạo.
Kiểu tuyến sinh dục giới tính (gonadal sex type) dựa vào sự hiện diện dịch hoàn hay buồng trứng.
1.4.2.2. Progesterone
Graham và Clarke (1997) ghi nhận progesterone vận chuyển trong máu dưới dạng liên kết với transcortin (một loại globulin liên kết với corticosteroid) ở nhiều loài động vật có vú. Trong dịch tử cung của thỏ, vào khoảng ngày thứ 3 đến ngày thứ 12 của thai kỳ, một chất vận chuyển progesterone được xác định là uteroglobin. Chất này có vai trò bảo vệ phôi thai trong suốt giai đoạn thú cái mang thai, nhưng cơ chế chưa rõ. Một loại protein trong huyết tương của chuột lang mang thai, chất này có vai trò vận chuyển progesterone, chất này được tổng hợp từ nhau thai và chịu sự kiểm soát của progesterone.
Progesterone có vai trò chính trong sinh sản của động vật cái, bao gồm sự xuất noãn, làm tổ của noãn thụ tinh và định vị của thai; trong đó, progesterone điều hoà chức năng của tử cung trong suốt chu kỳ động dục, không những ảnh hưởng đến quá trình làm tổ của noãn thụ tinh mà còn duy trì sự mang thai bằng cách kích thích sự phát triển các cấu tạo mô học ở tử cung và làm giảm co thắt của lớp cơ (Mani và ctv., 1997 ; Root Kustritz, 2001 ).
Fieni và ctv. (2001), Beijerink và ctv. (2003) ; Baan và ctv (2005) sử dụng các chất thuộc nhóm antiprogesterone như aglepristone hay bromocriptine để chấm dứt tình trạng mang thai, điều biến sự sinh đẻ trên chó, bởi vì chất này có cấu tạo tương đồng với progesterone nên tranh đoạt thụ thể với progesterone.
1.4.3. Sự biến thiên hàm lượng kích thích tố trong chu kỳ động dục
Sự biến thiên hàm lượng kích thích tố LH, estrogen, progesterone trong huyết thanh, sự sưng phồng âm hộ và quá trình sừng hoá các tế bào biểu mô âm đạo trong chu kỳ động dục của chó cái được Nelson (1998) diễn giải ở đồ thị 1.1.
Giai đoạn trước động dục: Köster và ctv. (2001) cho rằng cơ thể của chó cái chịu tác động của estrogen vào giai đoạn trước động dục. Estrogen được tổng hợp và phân tiết bởi các nang noãn phát triển, vì thế giai đoạn trước động dục được xếp vào pha estrogen hay pha nang noãn.
Vào cuối giai đoạn nghỉ ngơi, FSH và LH từ não thùy trước kích thích các nang noãn phát triển, thành thục, tổng hợp và phân tiết estrogen. Sang giai đoạn trước động dục, hàm lượng estrogen huyết thanh tăng dần sẽ làm thay đổi hành vi của chó cái, âm hộ thải dịch, ve vãn thú đực, chuẩn bị tử cung cho sự mang thai và một số thay đổi khác... Hàm lượng estrogen ở giai đoạn trước động dục có thể nâng lên >15 pg/ml. Cụ thể vào thời điểm đầu giai đoạn trước động dục thì lượng estrogen huyết thanh >25 pg/ml và gần cuối giai đoạn thì lượng estrogen huyết thanh đạt đến 60 - 70 pg/ml. Đỉnh điểm của estrogen đạt được vào lúc 24 - 48 giờ trước khi giai đoạn trước động dục chấm dứt và chó cái chuyển sang giai đoạn động dục (giai đoạn động dục đứng ì). Progesterone huyết thanh ở thời điểm 24 - 72 giờ cuối giai đoạn trước động dục rất thấp (< 0,5 ng/ml) (Concannon và ctv., 1975; Olson và ctv., 1984 ). Giai đoạn động dục Hàm lượng estrogen huyết thanh đạt trị số cực đại vào 1 - 2 ngày trước giai đoạn động dục và giảm dần trong giai đoạn động dục. Thực tế, chó cái thường biểu lộ các dấu hiệu động dục khi hàm lượng estrogen huyết thanh bắt đầu giảm, phản ảnh tiến trình nang noãn thành thục đã hoàn thành, vài ngày sau là hiện tượng xuất noãn và thành lập hoàng thể. Thực nghiệm chứng minh vai trò của estrogen đối với quá trình động dục trên 9 chó cái bị cắt bỏ buồng trứng khi được cung cấp estrogen. Các chó này có những dấu hiệu và hành vi của giai đoạn trước động dục, nhưng chúng không động dục (Johnston, 1980 - trích dẫn của Feldman và Nelson, 1996 Đồng thời, Feldman và Nelson (1996) cho rằng sự kết hợp giữa lượng progesterone huyết thanh tăng và estrogen huyết thanh giảm vào những ngày cuối của giai đoạn trước động dục đưa đến 2 kết quả: một là thay đổi hành vi và hai là tác động hồi phản dương đến vùng dưới đồi và tuyến não thùy, ảnh hưởng đến sự phân tiết FSH và LH. Vào giai đoạn trước động dục, hàm lượng progesterone huyết thanh chỉ đạt 0,5 - 1,0 ng/ml và >1,0 ng/ml trước khi xuất noãn. Khi LH đạt đỉnh, progesterone đạt 2 - 4 ng/ml; sau khi xuất noãn 2 ngày, progesterone nâng lên 4 - 10 ng/ml, do sự hình thành hoàng thể. Nồng độ progesterone huyết thanh ở giai đoạn động dục tiếp tục tăng và kéo dài trong nhiều tuần lễ khi chó cái bước sang giai đoạn sau động dục (Olson và ctv., 1984 ; Root Kustritz, 2000).
Giai đoạn sau động dục: Trong khoảng 5 - 20 ngày sau động dục, hàm lượng estrogen trong huyết thanh giảm dần đến khi đạt mức cân bằng như giai đoạn nghỉ ngơi. Ngược lại, hàm lượng progesterone ở giai đoạn này tiếp tục tăng cao và đạt tối đa vào khoảng 20 - 30 ngày sau khi xuất noãn, có thể lên đến 15 - 60 ng/ml và duy trì trong 2 - 3 tuần (Olson và ctv., 1984 ).
Thực nghiệm cho thấy chó mang thai có hàm lượng progesterone huyết thanh cao hơn chó cái không mang thai và kéo dài trong nhiều tuần từ khi không còn động dục; điều này rất quan trọng khi chẩn đoán mang thai bằng cách xét nghiệm progesterone (Johnston, 1980 - trích dẫn của Feldman và Nelson, 1996 ).
Pha hoàng thể ở chó không mang thai suy yếu dần, thường duy trì trong 10 - 30 ngày,nguyên nhân là do hoàng thể bị thoái hoá dưới tác động của prostaglandin. LH được sản xuất và phân tiết từ tuyến não thùy trước có vai trò trong sự thành lập hoàng thể nhưng không phải là yếu tố quan trọng, có thể prolactin là yếu tố quan trọng trong khoảng ½ thời gian sau khi hoàng thể được thành lập (Davol, 2000 [44]). Hàm lượng estrogen huyết thanh trong giai đoạn này cân bằng như ở giai đoạn nghỉ ngơi, mặc dù có sự phân tiết FSH và LH từng đợt của não thùy trước. Kích thích tố quan trọng trong giai đoạn sau động dục là prolactin. Hàm lượng prolactin trong huyết thanh thấp ở giai đoạn nghỉ ngơi, trước động dục và động dục nhưng đến khoảng giữa giai đoạn sau động dục thì nồng độ prolactin gia tăng, hiện tượng này xảy ra trên chó mang thai và không mang thai như nhau
Giai đoạn nghỉ ngơi: Tương tự như những loài động vật khác, ở vài cá thể, sự phân tiết LH vẫn xảy ra vào giai đoạn nghỉ ngơi. Giả thuyết cho rằng sự phân tiết kích thích tố của tuyến não thùy không phải tình cờ mà là cơ chế mồi cần thiết cho các nang noãn chưa phát triển của chu kỳ kế tiếp, các nang noãn này có tác động điều hoà quá trình phân tiết gonadotropin não thùy trước theo cơ chế hồi phản âm, ảnh hưởng đến sự động dục, xuất noãn và thụ tinh (Feldman và Nelson, 1996 ).
Nồng độ estrogen huyết thanh dao động trong suốt giai đoạn nghỉ ngơi, chỉ gia tăng vào giữa giai đoạn (20 - 46 pg/ml). Hành vi estrogen đã được khảo sát tương ứng với những đợt sóng phát triển của nang noãn, chính sự phát triển của nang noãn đã tạo ra sự gia tăng tối thiểu nồng độ estrogen huyết thanh, bởi vì các nang noãn này không bao giờ thành thục hoàn toàn và chúng sẽ bị thoái hoá. Ngược lại, nồng độ progesterone huyết thanh luôn luôn được duy trì ở mức độ thấp nhất trong suốt giai đoạn thú cái nghỉ ngơi
1.4.4. Sự thay đổi cấu tạo vi thể ở đường sinh dục trong chu kỳ động dục
Sự thay đổi kích thước các cấu tạo vi thể của đường sinh dục phụ thuộc vào hàm lượng kích thích tố buồng trứng (estrogen và progesterone) trong huyết thanh ở từng giai đoạn của chu kỳ động dục.
 |
| Cấu tạo vi thể lớp biểu mô âm đạo |
1.5. XÉT NGHIỆM TẾ BÀO ÂM ĐẠO
1.5.1. Sơ lược lịch sử
Theo dẫu liệu của Tirilomis và Malliarou (2003), phương pháp xét nghiệm tế bào biểu mô âm đạo còn gọi là xét nghiệm Pap (Pap test- Papanicolaou test) được Bác sỹ y khoa Papanicolaou (1833-1962) đề xướng vào năm 1928.
Ông bắt đầu nghiên cứu chu kỳ động dục của các loài động vật, sau đó nghiên cứu chu kỳ kinh nguyệt của phụ nữ bằng xét nghiệm vết phết âm đạo. Từ các kết quả khảo sát, ông nhận định: “Quá trình biểu hiện tính dục của động vật cái được lập đi lập lại trong một khoảng thời gian, có tính chất chu kỳ, như trường hợp chu kỳ kinh nguyệt ở người hay loài linh trưởng”.
Năm 1928, Papanicolaou báo cáo phương pháp xét nghiệm tế bào âm đạo dùng trong chẩn đoán ung thư đường sinh dục của phụ nữ. Ông bắt đầu quan tâm đến việc xét nghiệm các tế bào ung thư ở tử cung hoặc sừng tử cung bị bong tróc qua vết phết âm đạo và năm 1941, ông trình bày cách thu thập vết phết và phương pháp nhuộm các tế bào ung thư ở tử cung. Từ đó, xét nghiệm tế bào âm đạo được công nhận là phương pháp chẩn đoán y khoa, được áp dụng rộng rãi ở nhiều quốc gia trên thế giới và được gọi là xét nghiệm Pap.
1.5.2. Lớp biểu mô âm đạo
Lớp biểu mô âm đạo là lớp tế bào màng niêm lót mặt trong âm đạo, có nguồn gốc nội biểu mô (lá phôi trong), thuộc loại biểu mô phủ lát kép có thể hoá sừng .
Theo Concannon và De Gregario (1986) sự biến đổi ở lớp màng nhầy âm đạo là kết quả của quá trình gia tăng nồng độ estrogen trong huyết thanh của chó cái trong suốt giai đoạn trước động dục và kéo dài sang giai đoạn động dục. Sự biến đổi này phản ảnh qua các tế bào biểu mô âm đạo bị bong tróc. Vào giai đoạn trước động dục, nang noãn thành thục và tăng tiết estrogen. Estrogen làm lớp niêm mạc âm đạo tăng sinh, đẩy dần lớp tế bào ở ngoài xa dần mạch quản, chúng không được cung cấp máu nên chết dần và bong tróc. Hiện tượng chết của các tế bào biểu mô âm đạo được gọi là sự hoá sừng.
Để tìm hiểu ảnh hưởng của estrogen đến sự thay đổi cấu trúc của các tế bào biểu mô âm đạo trong quá trình sừng hoá. Cooper và ctv. (1966) tiến hành khảo sát trên nhóm chuột 10 ngày tuổi bằng cách tiêm 0,1 µg estradiol/chuột/ngày, trong 4 ngày liên tục. Kết quả ghi nhận lớp tế bào biểu mô âm đạo tăng sinh, nhiều lớp và hình dạng tế bào thay đổi từ hình khối dày, đến hình nhiều góc cạnh, cuối cùng tế bào biểu mô phẳng dẹp lại và hoá sừng. Ngoài ra, với kính hiển vi điện tử, các tác giả còn ghi nhận các tiểu thể trong tế bào biểu mô âm đạo cũng thay đổi trong quá trình sừng hoá. Ở giai đoạn đầu, trong các tế bào biểu mô nền gia tăng số lượng ri bô thể (ribosome), thể liên kết tế bào (desmosome) và hệ thống sợi trong tế bào chất (đường kính khoảng 70Å), nhân tế bào còn nguyên vẹn cấu trúc với màng nhân và nhiễm sắc thể trong dịch nhân. Ở tế bào biểu mô trung gian, các sợi trong tế bào chất gia tăng kích thước, đường kính lên đến 700 Å, ty thể và hệ thống lưới võng nội biến mất, các hạt keratohyaline xuất hiện, nhân tế bào bị thoái hoá dần. Ở tế bào biểu mô dẹp hay tế bào biểu mô chết với các sợi trong tế bào chất tập hợp lại, các hạt keratohyaline và nhân tế bào biến mất hoàn toàn.
Vijayasaradhi và Gupta (1987) khảo sát quá trình sừng hoá các tế bào biểu mô âm đạo của chuột và phân tích vai trò điều dẫn của estrogen được thực hiện ở in vitro. Kết quả ghi nhận các tế bào biểu mô âm đạo được nuôitrong môi trường dinh dưỡng và calcium vẫn phát triển, nhưng riềm lông tơ trên bề mặt tế bào biểu mô thì thưa thớt và cùn đi. Ngược lại, khi cung cấp thêm 17β estradiol, các tế bào biểu mô phát triển mạnh hơn, riềm lông tơ dầy đặn, tạo thành các hình chóp trên bề mặt tế bào. Các tác giả cho rằng chất 17β estradiol có vai trò điều dẫn tiến trình phát triển riềm lông tơ và các hình chóp vi tơ trên bề mặt tế bào là tính chất đặc hiệu của các tế bào biểu mô âm đạo bị hoá sừng. Đồng thời, Vijayasaradhi và Gupta (1988) ; Vijayalakshmi và Gupta (1997) xác định men transglutaminase phụ thuộc calcium có vai trò quan trọng trong quá trình sừng hoá, men transglutaminase xúc tác phản ứng phân giải polypeptide keratin tạo ra sản phẩm dipeptide γ-glutamyl-ε-lysine dưới sự kích hoạt của estradiol.
Tương tự, Gupta và ctv. (1989) chứng minh vai trò của 17 β estradiol trong quá trình sừng hoá các tế bào biểu mô âm đạo của chuột qua kính hiển vi điện tử truyền dẫn. Các tác giả ghi nhận hiện tượng các tế bào biểu mô âm đạo bong tróc và hoá sừng xảy ra vào giai đoạn động dục. Quá trình sừng hoá lớp biểu mô âm đạo xuất hiện sau khi tiêm 17 β estradiol 24 giờ trên nhóm chuột thí nghiệm, lớp biểu mô tăng sinh cao độ, tế bào biểu mô bong tróc thay đổi hình dạng, tế bào dẹp lại và trải rộng, sự liên kết giữa các tế bào trong lớp biểu mô không còn chắc chắn.
Ngoài ra, Chateau và Boehm (1996) còn xác định quá trình phân hoá của các tế bào bị hoá sừng ở lớp biểu mô âm đạo của chuột còn chịu sự kiểm soát của all-trans retinoic acid.
Rao và ctv. (1997) ghi nhận protein keratin bao gồm 4 loại, trọng lượng phân tử trung bình là 45, 47, 49 và 55 kDa; đồng thời, xác định estradiol có vai trò quan trọng đến tiến trình sinh tổng hợp keratin trong tế bào biểu mô âm đạo của chuột, estradiol tác động đến quá trình sao chép và giải mã của protein keratin.
Theo Higuchi và ctv. (2001) tác động xúc tác của men transglutaminase trong tế bào sinh keratin được kích hoạt bởi hợp chất sphigosylphosphorylcholine.
England và Concannon (2002) khảo sát lớp màng nhày âm đạo bằng kỹ thuật nội soi âm đạo. Các tác giả dựa vào nếp gấp và màu sắc lớp niêm mạc hoặc dịch chất trong âm đạo để xác định thời điểm phối chó cái. Ở giai đoạn trước động dục, các nếp gấp nở rộng, sưng phù, hồng nhạt với dịch nhầy lấp đầy các khe. Màu sắc của lớp nhày thay đổi dần từ màu hồng đậm sang hồng nhạt đến màu trắng kem phụ thuộc vào sự tăng sinh lớp tế bào biểu mô âm đạo . Vào cuối giai đoạn trước động dục và đầu động dục, các nếp gấp co rút lại và màu nhợt nhạt. Tình trạng này kéo dài trong khoảng 3 - 4 ngày khi nồng độ LH đạt đỉnh. Ở đầu giai đoạn sau động dục, khoảng 9 ngày sau khi hàm lượng LH đạt đỉnh, lớp màng nhầy âm đạo khô ráo và màu sắc đỏ hồng hơn.
Thí nghiệm theo dõi trên 3 chó cái ở giai đoạn nghỉ ngơi, 3 chó ở giai đoạn trước động dục và 3 chó ở giai đoạn động dục bằng kỹ thuật nhuộm hoá mô miễn dịch protein Ki 67, Steinhauer và ctv (2004) ghi nhận sự thay đổi hình thái lớp biểu mô ở đường sinh dục:
- Giai đoạn nghỉ ngơi, hàm lượng estrogen huyết thanh trung bình 19,8 pg/ml, progesterone 0,4 ng/ml, bề dày lớp tế bào biểu mô không có lông rung là 7,85 μm và lớp tế bào biểu mô có lông rung là 12,52 μm.
- Vào giai đoạn trước động dục và cuối pha nang noãn, estrogen huyết thanh tăng lên 41,8 pg/ml, progesterone 2,9 ng/ml, tương ứng với bề dày lớp tế bào biểu mô không có lông rung là 25,82 μm và tế bào biểu mô có lông rung là 23,53 μm.
- Giai đoạn động dục (giữa pha hoàng thể), estrogen đạt 36,6 pg/ml và progesterone đạt 29,6 ng/ml; chiều cao các tế bào biểu mô không có lông rung là 13,67 μm và tế bào biểu mô có lông rung 13,38 μm.
Điều đó cho thấy, sự tăng sinh lớp tế bào biểu mô ở đường sinh dục chó cái phụ thuộc vào cung lượng estrogen huyết thanh và tăng sinh cao nhất vào giai đoạn trước động dục (pha nang noãn). Do vậy, phân tích tế bào âm đạo có thể xem như là xét nghiệm gián tiếp hàm lượng estrogen huyết thanh.
 |
| Cấu tạo vi thể của sừng tử cung Ghi chú: BMTC. lớp biểu mô sừng tử cung CƠTC. lớp cơ sừng tử cung NTTC. nang tuyến sừng tử cung MLK. mô liên kết sừng tử cung |
1.5.3. Sự biến đổi các loại tế bào biểu mô âm đạo trong chu kỳ động dục
1.5.3.1. Các loại tế bào biểu mô âm đạo
Hopkins (2003) ghi nhận estrogen được phân tiết từ nang noãn có tác động đến sự phát triển và thành thục các tế bào biểu mô âm đạo. Do vậy các tế bào biểu mô sẽ chuyển đổi hình dạng từ tế bào nền sang cận nền, đến trung gian và tế bào bề mặt với số lượng khác nhau tương ứng ở từng thời điểm của từng giai đoạn trong chu kỳ động dục. Dựa vào sự biến đổi này, xét nghiệm tế bào âm đạo có thể được sử dụng để giám sát chu kỳ động dục của chó cái, đặc biệt giai đoạn trước động dục và động dục là hai giai đoạn điển hình.
Tế bào biểu mô âm đạo gồm tế bào cận nền, tế bào trung gian nhỏ, tế bào trung gian lớn (trung gian-bề mặt) và tế bào bề mặt.
Tế bào cận nền là tế bào biểu mô có kích thước nhỏ nhất trong các loại, tế bào hình tròn hoặc bầu dục, nhân rất to và tròn đều, tỉ lệ nhân/tế bào chất thấp. Lớp tế bào cận nền nằm trong cùng, gần lớp tế bào mầm (tế bào nền), được cung cấp máu đầy đủ.
Tế bào trung gian có kích thước rất thay đổi, thường lớn hơn tế bào cận nền, nhưng nhân nhỏ hơn rất nhiều, hình dạng có góc cạnh. Sự thay đổi hình thái tế bào biểu mô từ dạng cận nền chuyển sang trung gian là giai đoạn mở đầu cho sự chết của tế bào biểu mô âm đạo. Tế bào trung gian có thể phân làm 2 loại, tế bào trung gian nhỏ và tế bào trung gian lớn.
Tế bào trung gian-bề mặt là tế bào biểu mô âm đạo chết điển hình, chúng xuất hiện trong vết phết âm đạo của chó cái vào cuối giai đoạn trước động dục, tế bào có nhiều góc cạnh, dẹp, vách tế bào không còn rõ ràng, tế bào chất và nhân bị tiêu hủy, vì thế nhân rất nhỏ. Eilts (2001) gọi tế bào này là tế bào bề mặt.
Tế bào bề mặt hay tế bào vảy, là các tế bào biểu mô chết hoàn toàn, không còn nhân, hình dạng dẹp và phẳng; vì thếtrong vết phết, các tế bào bề mặt thường bị gấp nếp và hiện diện vào giai đoạn chó cái động dục. Eilts (2001) gọi tế bào này là tế bào không nhân. Ngoài ra, tác giả chia các tế bào biểu mô trong vết phết âm đạo dựa vào sự sừng hoá thành 2 nhóm: nhóm tế bào biểu mô không sừng hoá (tế bào cận nền và trung gian) và nhóm tế bào sừng hoá (tế bào bề mặt và tế bào vảy không nhân). Vì thế, hình tượng tháp nghiêng PISA ở Ý biểu thị quá trình sừng hoá của tế bào biểu mô.
1.5.3.2. Tỉ lệ các loại tế bào biểu mô âm đạo trong chu kỳ động dục Các loại tế bào biểu mô âm đạo ở giai đoạn trước động dục
Cibas (2002) ; Spano (2002) ; Beimborn và ctv. (2003) khảo sát vết phết âm đạo của chó cái ở giai đoạn trước động dục và ghi nhận tế bào cận nền, trung gian và một vài tế bào bề mặt bị bong tróc; ngoài ra còn có các tế bào khác như hồng cầu, bạch cầu và vi khuẩn.
Tuy nhiên, Feldman và Nelson (1996) ghi nhận: vào đầu giai đoạn trước động dục, các loại tế bào biểu mô âm đạo tương tự như cuối giai đoạn nghỉ ngơi, chủ yếu gồm tế bào cận nền và tế bào trung gian, nhưng có thể phân biệt hai giai đoạn bằng cách dựa vào sự hiện diện của hồng cầu, bạch cầu trung tính và vi khuẩn. Giữa giai đoạn trước động dục, tiêu bản vết phết âm đạo cho thấy bạch cầu trung tính biến mất, nhiều hồng cầu và tỉ lệ tế bào bề mặt gia tăng, thay thế dần các tế bào trung gian và trung gian-bề mặt. Cuối giai đoạn trước động dục, vết phết có thể có một ít hồng cầu, không có bạch cầu; vì thế, tiêu bản trở nên trong sáng và dễ quan sát hơn. Tế bào bề mặt chiếm tỉ lệ cao. Ở một vài cá thể, vết phết âm đạo vào cuối giai đoạn vẫn còn xuất hiện nhiều hồng cầu, tương ứng với dịch xuất ở âm hộ có lẫn máu.
Hơn nữa, thời gian trước động dục của chó cái có thể thay đổi từ 2 - 21 ngày và vết phết âm đạo vào cuối giai đoạn trước động dục có tỉ lệ tế bào bề mặt chiếm > 80% và tỉ lệ tế bào biểu mô không còn nhân có thể 100% Các loại tế bào biểu mô âm đạo ở giai đoạn động dục. 90% chúng hiện diện ³Hopkins (2003) ghi nhận vết phết âm đạo chó cái ở giai đoạn động dục có số tế bào bề mặt trong suốt giai đoạn động dục, không có tế bào cận nền và trung gian; ngoài ra, hồng cầu và vi khuẩn có thể có hoặc không. Nếu bạch cầu hiện diện với số lượng nhiều, chứng tỏ chó cái bị viêm nhiễm đường sinh dục.
Theo Feldman và Nelson (1996) trong giai đoạn chó cái động dục, tế bào bề mặt không nhân (tế bào vảy) chiếm 100 %, tương ứng với giai đoạn chó cái “động dục đứng ì”, hàm lượng LH huyết thanh đạt đỉnh, sự xuất noãn xảy ra và đây cũng là thời điểm phối chó cái ...Hơn nữa, sự bong tróc các tế bào biểu mô gia tăng, là kết quả phản ảnh của sự gia tăng hàm lượng estrogen huyết thanh ở giai đoạn trước động dục.
Các loại tế bào biểu mô âm đạo ở giai đoạn sau động dục
Nếu thú cái không được phối hoặc phối không đậu thai sẽ chuyển từ giai đoạn động dục sang giai đoạn sau động dục.
Dore (1978) ; Feldman và Nelson (1996) ghi nhận các loại tế bào biểu mô âm đạo của chó cái ở vào giai đoạn sau động dục có khác biệt rõ so với giai đoạn động dục. Vào giai đoạn động dục, tế bào bề mặt chiếm khoảng 100%, sau 24 - 48 giờ chấm dứt giai đoạn động dục, tế bào bề mặt còn khoảng 20%, chúng được thay thế bằng tế bào cận nền và trung gian.
Ngoài ra,trong tiêu bản vết phết, bạch cầu trung tính có thể xuất hiện trở lại, nhưng thường không nguyên vẹn hoặc nằm trong tế bào chất của tế bào biểu mô, tế bào này được gọi là “tế bào không động dục” (metestrum cell). Dựa vào sự hiện diện của “tế bào không động dục” và hồng cầu giúp chúng ta có thể phân biệt chó cái ở giai đoạn sau động dục hay đầu giai đoạn trước động dục
Vào khoảng giữa của giai đoạn sau động dục, tập đoàn tế bào biểu mô trong vết phết âm đạo chủ yếu là tế bào cận nền, tương tự như vết phết âm đạo ở giai đoạn nghỉ ngơi Các loại tế bào biểu mô âm đạo ở giai đoạn nghỉ ngơi
Trong giai đoạn nghỉ ngơi, chó cái không thể hiện dục tính, cơ quan sinh dục phục hồi và sửa chữa lớp niêm mạc đường sinh dục. Vì thế, các tế bào biểu mô âm đạo bong tróc rất ít, vết phết âm đạo được ghi nhận chủ yếu là tế bào cận nền và tế bào trung gian, có thể có vài tế bào bạch cầu và vi khuẩn. ./.
Receive articles via Email!